Бактерии представляют собой одноклеточные организмы, лишенные хлорофилла. Эта группа микроорганизмов наиболее многочисленна, широко распространена в природе и хорошо изучена. Среди бактерий имеется значительное число возбудителей инфекционных заболеваний человека и животных.
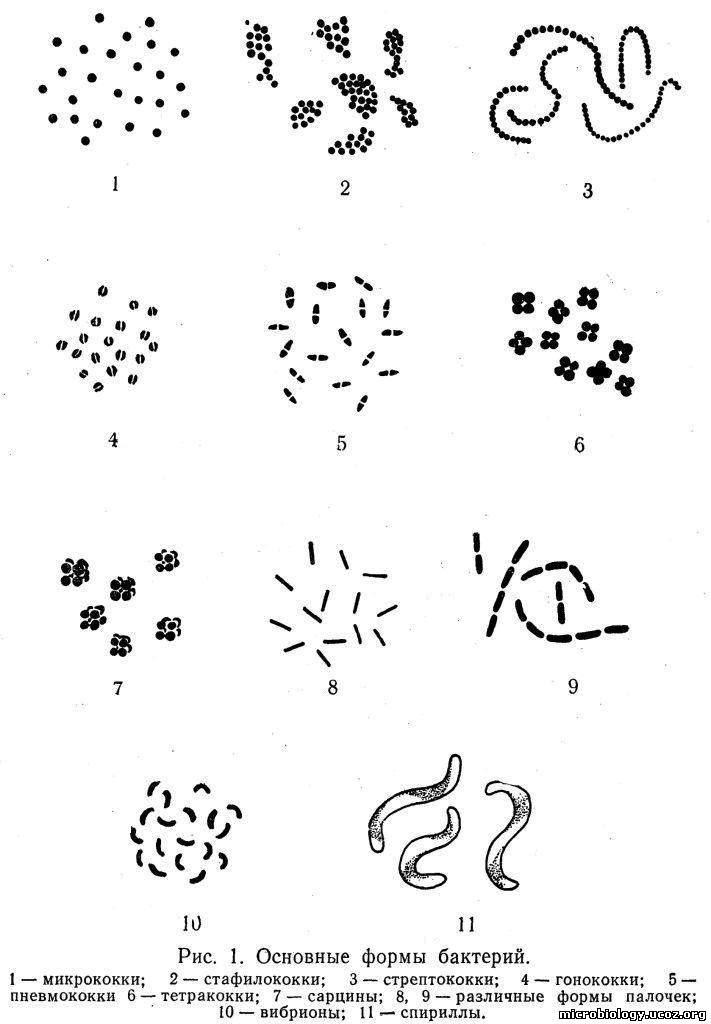
Форма и размеры бактерий. По форме клеток бактерии разделяются на шаровидные — кокки; палочковидные или цилиндрические — собственно бактерии; извитые — вибрионы и спириллы. Между основными формами имеются переходные. Различные формы бактерий показаны на рис. 1.
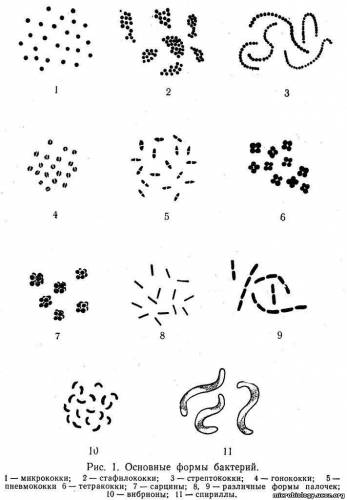
Кокки (от греч. coccus — зерно, ягода) различаются между собой в зависимости от расположения клеток после их деления. Одиночные кокки называются микрококками (рис. 1,1), парные — диплококками. Если кокки после деления не расходятся, а образуют цепочку, их называют стрептококками (рис. 1,3). Все эти кокки делятся только в одной плоскости. При делении в двух взаимно перпендикулярных плоскостях могут образоваться сочетания из четырех кокков — тетракокки (рис. 1,6), а при делении в трех взаимно перпендикулярных плоскостях — сарцины (от лат. sarcio — связывать; рис. 1,7), состоящие из 8 — 16 клеток. Если деление происходит без определенного порядка, кокки остаются вместе и образуют скопления, напоминающие грозди винограда, — стафилококки (рис.1,2). Обычно размеры кокков достигают 1 — 1,5 мкм.
Среди кокков имеются возбудители различных заболеваний человека: диплококки-пневмококки (рис. 1,5), мейингококки и гонококки (рис. 1,4) вызывают соответственно воспаление легких, менингит и гонорею; стафилококки и стрептококки — различные гнойные заболевания человека и животных. Многие кокки являются обитателями различных полостей и кожи человека и широко распространены во внешней среде.
Палочковидные бактерии (от греч. bacteria — палочка) имеют цилиндрическую форму и обычно располагаются одиночно (рис. 1,8 — 9), но иногда попарно (диплобактерии) или в виде цепочек (стрептобактерии). Палочки могут быть прямыми, слегка изогнутыми и веретенообразными; размеры их достигают 1 — 5x0,5 — 1 мкм. Палочки, не образующие спор, называют бактериями, а спорообразующие — бациллами (аэробы) и клостридиями (анаэробы). Под воздействием различных факторов форма и величина бактерий могут меняться. Способность бактерий изменять свою форму и величину называется полиморфизмом.
Среди бактерий много возбудителей инфекционных заболеваний: чумы, сибирской язвы, бруцеллеза, столбняка, газовой гангрены, дифтерии, кишечных инфекций.
Извитые формы бактерий имеют вид спирали, состоящей из нескольких завитков. Среди них различают вибрионы, имеющие один завиток (рис. 1, 10), и спириллы с 2 — 3 завитками (рис. 1, 11).
Вибрионы — слабоизогнутые клетки, напоминающие запятую, длиной 1 — 3 мкм, очень подвижные за счет жгутика, расположенного на конце клетки. Среди вибрионов наибольшее значение имеет возбудитель холеры.
Спириллы — безвредные микроорганизмы, живущие в сточных или загрязненных водах, гниющих отбросах. Только Spirillum minus вызывает у человека болезнь укуса крысы — содоку.
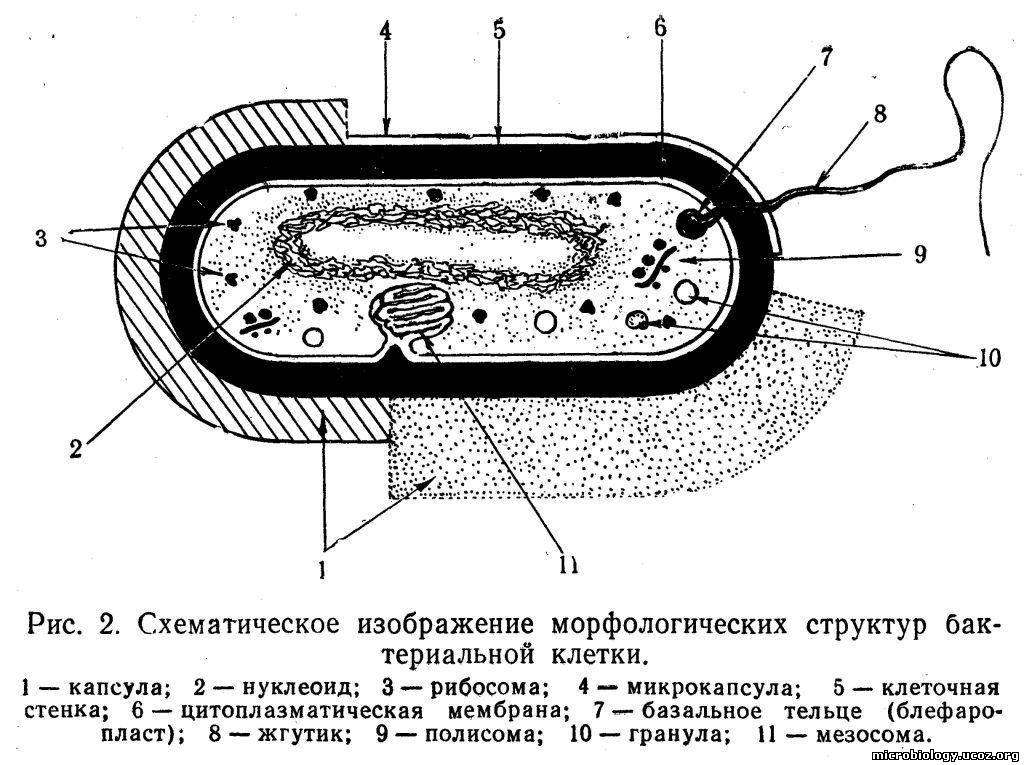
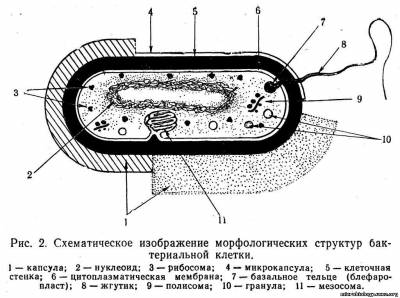
Структура бактерий. Бактериальная клетка состоит из клеточной стенки, цитоплазматической мембраны и цитоплазмы, которая содержит ядерное вещество, различные органеллы и включения. Кроме того, у многих бактерий имеются капсула и слизистый слой, жгутики и пили (рис. 2).
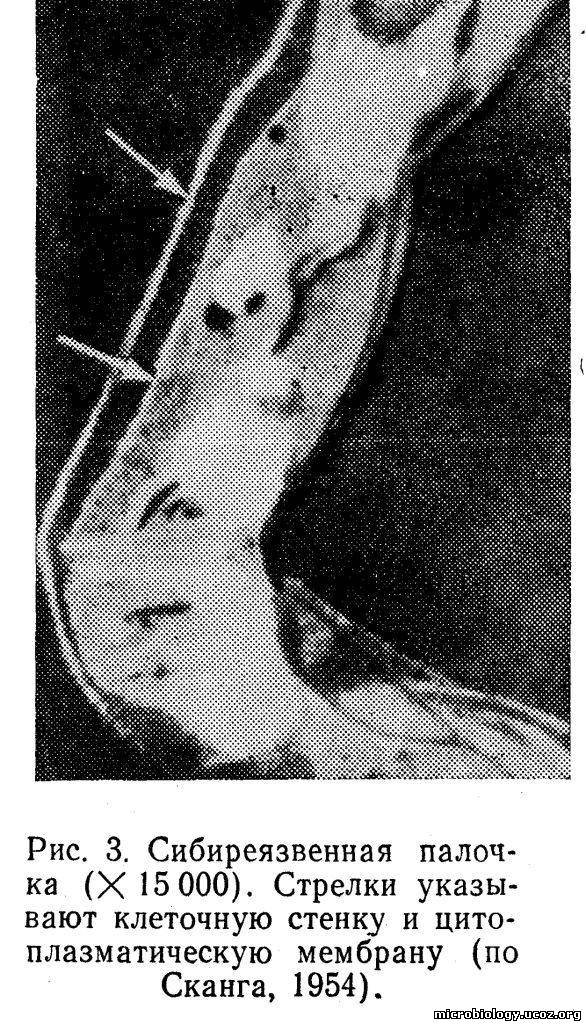
Клеточная стенка. Оболочка, которая отделяет микробную клетку от окружающей среды, определяет и сохраняет ее форму, получила название клеточной стенки (рис. 3). Она характеризуется прочностью, эластичностью и гибкостью. Клеточная стенка выполняет жизненно важную функцию: предохраняет клетку от осмотического лизиса, так как давление внутри клетки в цитоплазме выше, чем в окружающей среде. Обладая избирательной проницаемостью, клеточная стенка обеспечивает прохождение внутрь клетки различных веществ и выведение наружу продуктов обмена. Через клеточную стенку легко проникают вода, глюкоза, аминокислоты, жирные кислоты, имеющие молекулы небольших размеров. Более крупные молекулы органических веществ не могут проникнуть внутрь клетки без предварительного расщепления их на более мелкие с помощью ферментов, выделяемых клеткой.
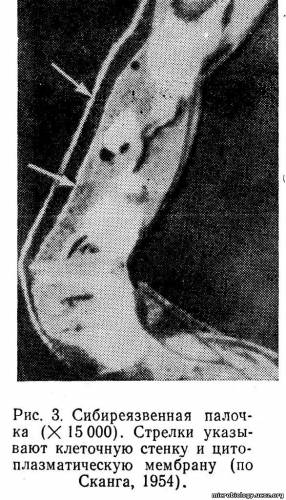
Клеточная стенка бактерий имеет сложную структуру и построена из компонентов двух типов. Прочность и твердость клеточной стенке придает сеть микрофибрилл, которая погружена в содержимое — матрикс. Микрофибриллы являются гликопептидами (пептидогликаны, или муреины). Слой гликопептидов определяет и сохраняет форму бактериальной клетки. Структура и химический состав клеточных стенок грамположительных и грамотрицательных бактерий различны.
Клеточная стенка грамположительных бактерий имеет наиболее простое строение. Структура ее однородна, она толще (10 — 15 нм), чем клеточная стенка грамотрицательных бактерий. Основная масса клеточной стенки — гликопептиды (до 90%). Сеть микрофибрилл погружена в матрикс, содержащий полисахариды (до 90%) и тейхоевые кислоты. Белки обычно отсутствуют, а липиды составляют всего 2,5%. Однако некоторые грамположительные бактерии, например коринебактерии и микобактерии, содержат в клеточной стенке большое количество липидов.
Клеточная стенка грамотрицательных бактерий имеет сложное строение и по химическому составу значительно отличается от клеточных стенок грамположительных бактерий. Внутренний слой клеточной стенки — тонкий мешочек молекул гликопептида, состоящий из одного или двух молекулярных слоев (2 — 3 нм). Поверх него лежит широкий внешний слой (7—8 нм) из неплотно упакованных молекул белка и фосфолипидов, над которым располагается третий слой — липополисахариды. Возможна и другая структура внешнего слоя клеточной стенки: в двойной слой фосфолипидов включены белки и липополисахариды.
В клеточной стенке этих бактерий много липидов (до 25%), белка и полисахаридов.
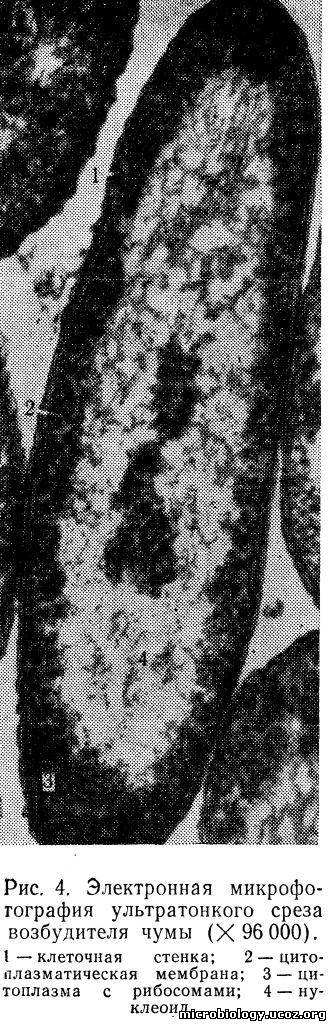
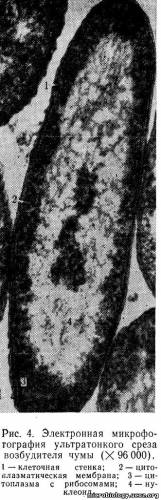
Цитоплазматическая мембрана. Непосредственно под клеточной стенкой расположена цитоплазматическая мембрана, очень плотно прилегающая к ней (рис.4). Цитоплазматическая мембрана имеет большое значение в жизни клетки. Она действует как осмотический барьер, концентрируя внутри клетки питательные вещества и способствуя выведению продуктов обмена. Через нее проходят частицы, имеющие молекулы небольших размеров (фрагменты ДНК, белки с низкой молекулярной массой — внеклеточные ферменты). Белки цитоплазматической мембраны — пермеазы выполняют функцию транспорта — переноса органических и неорганических веществ в клетку. Дитоплазматическая мембрана является местом биосинтеза некоторых составных частей клетки, принимает участие в процессах деления бактерий. На внутренней поверхности ее находятся специальные участки, к которым прикрепляется ДНК в процессе ее удвоения (репликации). Рост мембраны обеспечивает разделение генома клетки после завершения процесса репликации. У аэробных бактерий в цитоплазматической мембране находится цепочка переноса электронов, обеспечивающих энергетический обмен клетки.
Цитоплазматическая мембрана очень тонка (не более 8—10 нм). На электронных микрофотографиях она видна как двойная линия, разделенная светлым промежутком (трехслойная). Более половины массы цитоплазматической мембраны составляют белки и 20—30% — фосфолипиды. Цитоплазматическая мембрана бактерий имеет структуру элементарной биологической мембраны — двойного слоя фосфолипидов, на поверхности которых расположены белки.
При некоторых воздействиях на бактериальную клетку, например при помещении ее в гипертонический раствор хлорида натрия, мембрана может отделиться от клеточной стенки и стать хорошо видимой (см. рис. 3).
Цитоплазма. Содержимое бактериальной клетки — ограниченное цитоплазматической мембраной прозрачное, слегка вязкое вещество жидкой консистенции. Цитоплазма клеток бактерий является коллоидальной системой, состоящей из воды, протеинов, жиров, углеводов, различных минеральных и других веществ, соотношения которых варьируют в зависимости от вида бактерий и возраста клетки.
В цитоплазме бактерии находятся ядро клетки — нуклеоид, рибосомы, мезосомы, а также различные гранулы запасных питательных веществ, пигменты, жиры.
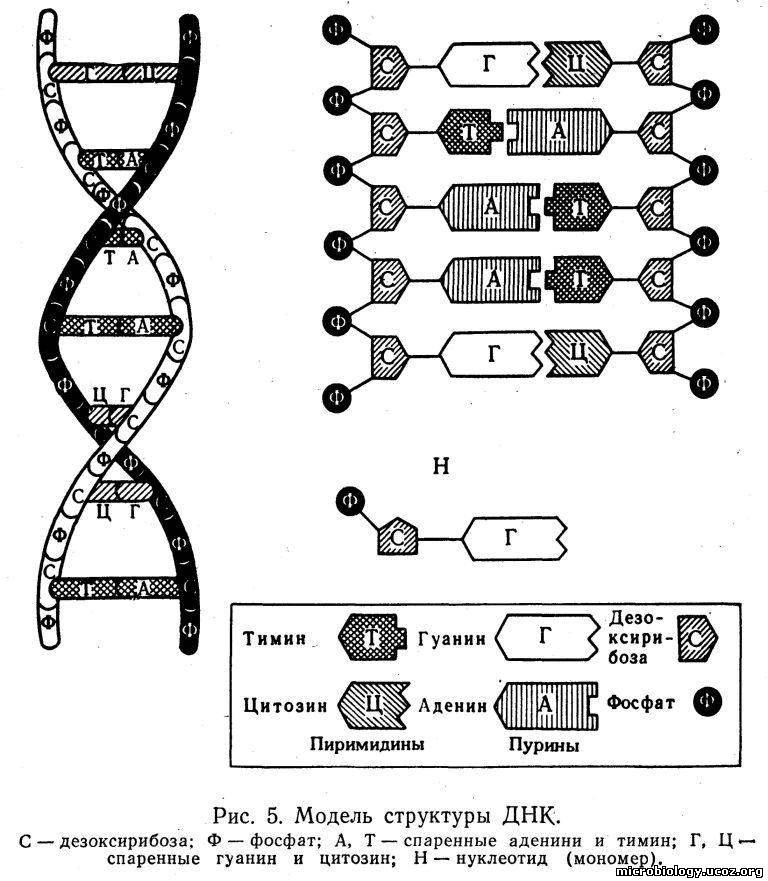
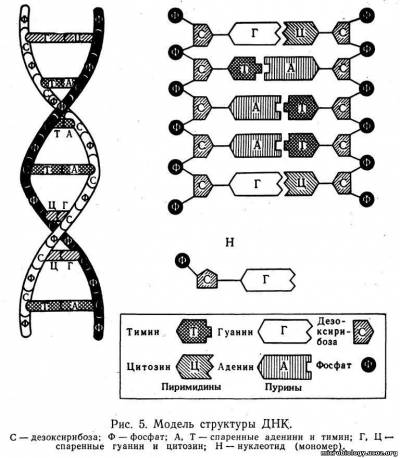
Нуклеоид. Содержит ДНК, которая связана с небольшим количеством специфического основного белка — гистона (нуклеопротеид) и является хранителем наследственной информации в клетке. В отличие от ядер других микроорганизмов, например простейших, нуклеоид бактерий не имеет ясно выраженной мембраны, ограничивающей его от остальной части цитоплазмы (см. рис. 4). Молекула ДНК по схеме, предложенной в 1953 г. Уотсоном и Криком, состоит из двух полинуклеотидных цепей, закрученных одна вокруг другой наподобие винтовой лестницы (рис. 5). Наружную поверхность такой двойной спирали образует сахар — дезоксирибоза (С), которая чередуется с остатками фосфорной кислоты (Ф). Внутри спирали перпендикулярно к ее оси, как ступеньки лестницы, расположены плоские молекулы азотистых оснований: пурины — аденин (А), гуанин (Г) и пиримидины — тимин (Т), цитозин (Ц). Каждый пурин вследствие своей химической структуры обязательно соединен с пиримидином, поэтому нить ДНК имеет равномерную толщину, около 0,2 нм, на всем протяжении. Длина молекулы ДНК может быть в сотни миллионов раз больше. Например, общая длина хромосомы кишечной палочки 1 — 1,4 мм. Пурины и пиримидины соединены между собой водородными связями, которые легко разрываются. Каждое азотистое основание прикреплено только к сахару наружной цепи — дезоксирибозе. Дезоксирибоза, фосфат и азотистое основание образуют один мономер ДНК, называемый нуклеотидом (Н). Для ДНК многих бактерий характерна кольцевая структура в виде замкнутого кольца. У большинства прокариотов только одна бактериальная хромосома.
Рибосомы. Помимо ДНК, в клетке есть вторая нуклеиновая кислота — рибонуклеиновая (РНК), которая в отличие от ДНК состоит из одной цепи, имеет сахар рибозу вместо дезоксирибозы и урацил вместо тимина. Основная масса РНК связана с белком в форме маленьких частиц, или рибосом, которые являются центрами синтеза белка. Рибосомы образуют большие агрегаты, называемые полирибосомами, или полисомами, состоящими из 7 — 8 рибосом и более. Химический состав рибосом: 40 — 60% РНК и 60 — 40% белка. У бактерий рибосомы свободно лежат в цитоплазме. Количество их в каждой клетке может быть более 100. Помимо рибосомальной РНК (рРНК), в цитоплазме бактерии находится еще информационная РНК (иРНК, или мРНК). Она осуществляет функцию переноса генетической информации от ДНК к полисомам. У кишечной палочки она составляет 2 — 4 % от всей РНК. Третья рибонуклеиновая кислота — транспортная (тРНК) — выполняет функцию транспортировки в рибосомы аминокислот, необходимых для синтеза белка.
Мезосомы. У некоторых бацилл из цитоплазматической мембраны возникают сферические, закрученные в завиток структуры — так называемые мезосомы. Функция их пока не совсем ясна. Возможно, они участвуют в процессе деления клетки или в окислительно-восстановительных процессах, выполняя роль митохондрий.
Гранулы. В цитоплазме бактерий находятся различные гранулы, многие из которых содержат запасные питательные вещества. Источником углерода или энергии служат гранулы безазотистых органических веществ — полисахариды, состоящие из молекул глюкозы. Одни гранулы состоят из крахмала и окрашиваются йодом в синий цвет (иогены или гранулеза), другие содержат гликоген и окрашиваются йодом в красновато-коричневый цвет. Сернистые бактерии накапливают в цитоплазме капельки серы, некоторые бактерии синтезируют и накапливают липидные включения, которые видны в форме мелких капель благодаря большой степени их преломления.
У некоторых микробов в цитоплазме находятся зерна волютина, впервые обнаруженные у спирилл (Spirillum volutans). Они являются запасными питательными веществами, состоящими из неорганических полифосфатов и соединений, близких к нуклеиновым кислотам. Волютин в виде крупных гранул накапливается в цитоплазме бактерий при выращивании их на средах, содержащих углеводы. Зерна волютина при окраске их метиленовым синим обнаруживают явления метахромазии: синяя краска придает им ярко-красный цвет. У некоторых бактерий, например коринебактерий, обнаружение зерен волютина является ценным диагностическим признаком.
Капсула и слизистый слой. У многих бактерий с наружной стороны клеточной стенки расположен диффузный гомогенный слизистый слой различной толщины (см. рис. 2,1). Этот слой можно выявить при определенных способах окраски или соответствующем освещении.
Капсулой называют слой, который сохраняет тесную связь с клеточной стенкой и служит внешним покровом клетки. Толщина его ограничена, и капсула четко выявляется при негативном окрашивании по методу Гинса: на темном фоне препарата видна окрашенная в красный цвет бактериальная клетка, окруженная бесцветной капсулой. Толщина капсул у бактерий различна: от долей микрометра до 10 мкм. Капсулу величиной менее 0,2 мкм часто называют микрокапсулой. Поверхностные структуры типа капсул описаны у пневмококков, возбудителей сибирской язвы, коклюша, гонореи, группы капсульных бактерий — клебсиелл. У многих видов бактерий капсула появляется лишь при определенных условиях, часто неблагоприятных. Возбудители сибирской язвы, коклюша, гонореи, пневмококки образуют капсулу, попадая в организм человека или животного. В этом случае капсула выполняет защитную роль, предохраняя микроб от действия антител, фагоцитов и других защитных факторов организма. Группа капсульных бактерий сохраняет капсулу постоянно: и в организме человека, и при культивировании на питательных средах. Химический состав капсул зависит от вида бактерий. Основными компонентами капсулы являются вода (до 98%) и полисахариды. В капсуле сибиреязвенных бацилл найдены полипептиды, а в капсуле стрептококка — белок М.
Слизистые слои, образующиеся вокруг поверхности некоторых бактерий, отличаются от капсул более рыхлым строением, толщиной, способностью частично отделяться от образовавшей их клетки. Материал, составляющий слизистый слой, часто обнаруживают в питательной среде, в которой культивируют микроорганизмы.
Защитные функции капсулы разнообразны. Помимо предохранения микроба от действия защитных факторов макроорганизма, капсула предохраняет микроб от притока в клетку большого количества жидкости (осмотический барьер), а также от высыхания при неблагоприятных условиях среды обитания.
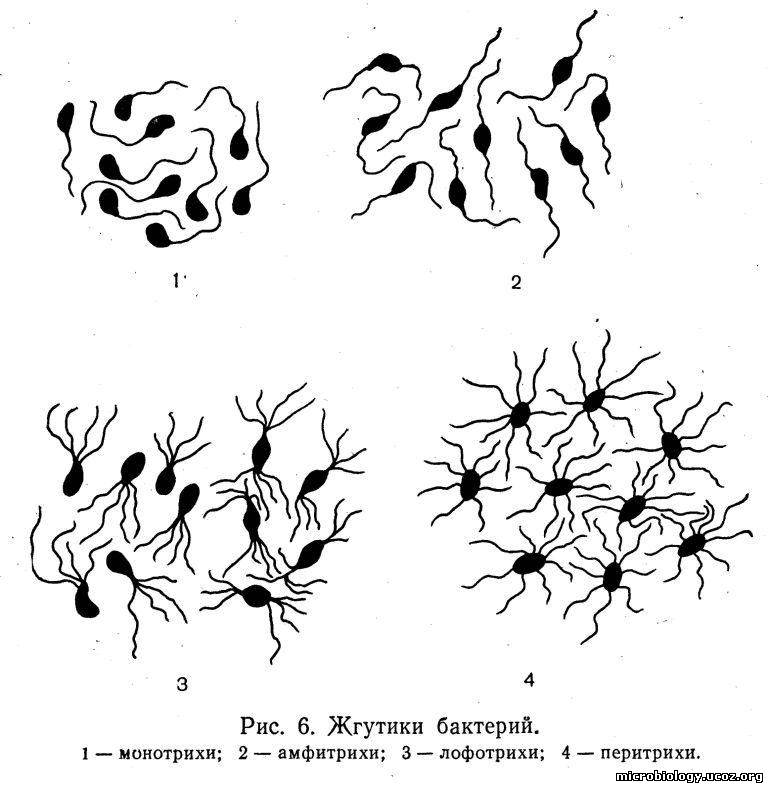
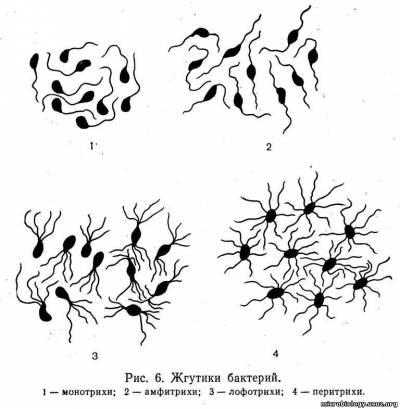
Жгутики. Некоторые бактерии обладают подвижностью, которая осуществляется с помощью жгутиков. Число и расположение жгутиков являются характерным видовым признаком бактерий, который используют для дифференциации микроорганизмов. По расположению и числу жгутиков различают бактерии: монотрихи, имеющие один жгутик на одном из полюсов клетки; амфитрихи, у которых на каждом полюсе расположено по одному жгутику; лофотрихи — с пучком жгутиков на одном полюсе (сюда же относят бактерии, которые имеют пучки жгутиков на обоих полюсах), и перитрих и, жгутики у которых расположены по всей поверхности тела (рис. 6).
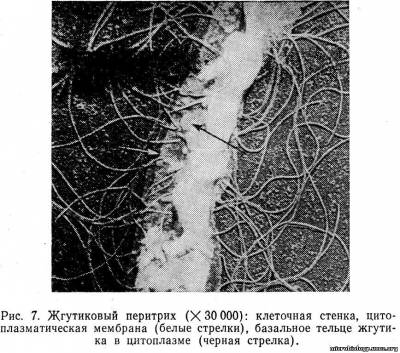
Жгутики представляют собой тонкие, спиральные, нитевидные фибриллы толщиной 12 — 18 нм. Длина жгутика может в 10 раз превышать длину самой бактерии. Жгутик отходит от специального образования — базального тельца, расположенного в цитоплазме на внутренней поверхности цитоплазматической мембраны (рис. 7). Базальное тельце имеет сложное строение, в нем находится механизм в виде двух кольцевых пластинок, вращение которых относительно друг друга сообщает движение жгутику.
Жгутики бактерий — белковые нити, состоящие из белка флагеллина, белковые мономеры которого собраны в спиральные цепи, закрученные вокруг полой сердцевины. При движении жгутик вращается вокруг своей длинной оси по или против часовой стрелки. Движение бактерий можно увидеть при исследовании их в живом состоянии с помощью метода висячей или раздавленной капли и при использовании специальных способов окраски в световом микроскопе. Скорость активного движения с помощью жгутиков у некоторых бактерий очень велика: за 1 с они могут пересечь расстояние, в 20 раз превышающее их длину. Механическое удаление приводит к потере подвижности бактерий, но не препятствует их росту и размножению.
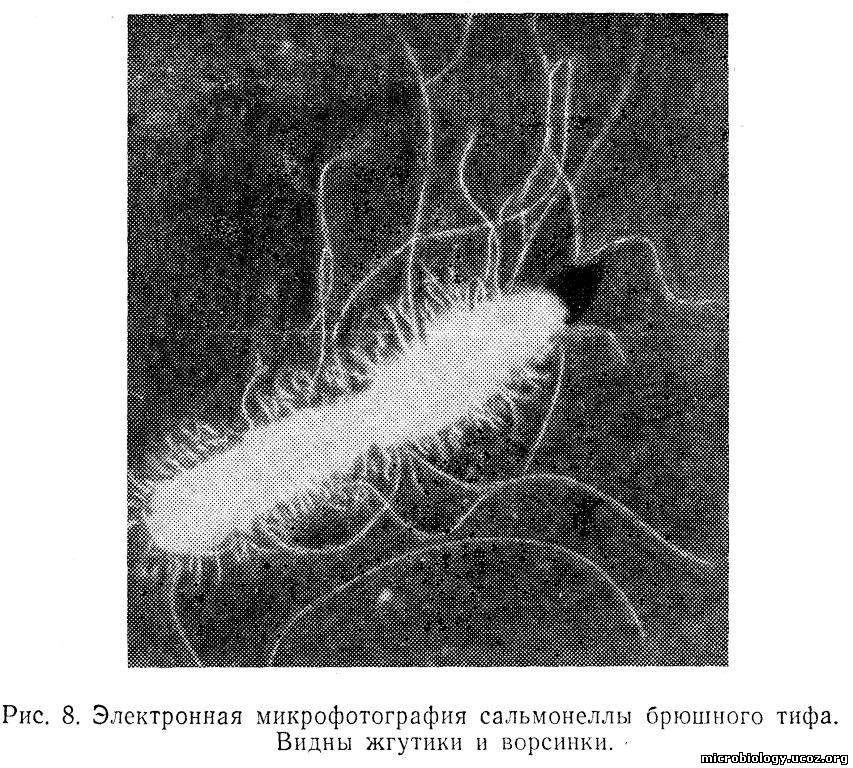
Пили (ворсинки). Прямые нитевидные образования, обнаруженные у сальмонелл, эшерихий, протея, называют ворсинками, а также бахромками, фимбриями, ресничками, пилями (рис. 8). Пили тоньше жгутиков бактерий и короче их; состоят из особого белка пилина, мономеры которого, как и у жгутиков, расположены по спирали. Пили различаются по диаметру и длине; толщина пилёй может быть от 4 — 10 до 35 нм. Количество пил ей на одну бактериальную клетку может достигать нескольких сотен. Пили обеспечивают способность бактерий к прилипанию (адгезия) друг к другу или к субстрату, например к эпителиальным клеткам слизистой оболочки кишечника.
Некоторые пили, например F-ворсинки, выполняют половые функции у бактерий. Они обеспечивают передачу наследственного материала (ДНК) из одной бактериальной клетки в другую, образуя мостик между двумя клетками. Эти ворсинки шире и длиннее остальных и на конце имеют шаровидное утолщение.
Споры. Некоторые бактерии, попадая в неблагоприятные условия существования, образуют внутри тела спору (эндоспора). Эндоспора представляет собой внутриклеточное, сильно преломляющее свет образование, устойчивое (резистентное) к различным вредным факторам внешней среды: высыханию, действию высоких температур, химических и дезинфицирующих веществ (рис. 9).
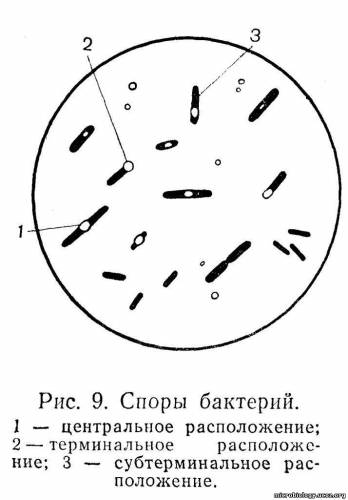
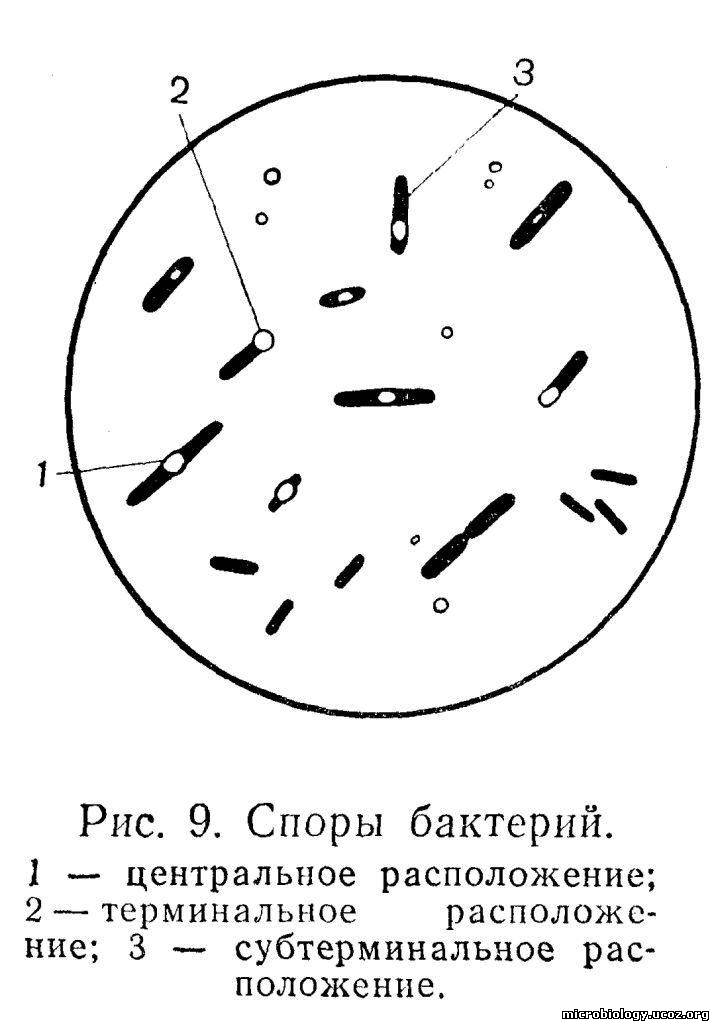
Спорообразование свойственно преимущественно палочковидным формам бактерий: бациллам и клостридиям. У бактерий других видов оно встречается очень редко. Споры имеют сферическую, овальную или эллипсоидную форму. Диаметр споры обычно равен диаметру клетки, в которой она образуется, или несколько превышает его, а длина споры составляет 1/4 -1/3 длины клетки бактерии. Размер и положение внутри бактериальной клетки зависят от вида, возраста и условий выращивания бактерий. Споры могут располагаться в центре клетки — центрально (рис. 9,1), как, например, у возбудителя сибирской язвы; ближе к концу — субтерминально, у возбудителя газовой гангрены (рис. 9,3); на самом конце — терминально, у возбудителя столбняка и ботулизма (рис. 9,2). Форма и расположение споры в бактериальной клетке могут быть отличительными признаками некоторых возбудителей: например, столбнячная палочка имеет круглую спору, расположенную на конце бактерии, и похожа на барабанную палочку, а ботулиническая палочка — овальную спору также на конце бактериальной клетки и напоминает теннисную ракетку. Созревшая спора имеет сложную структуру.
Процесс спорообразования происходит при попадании бактерии в неблагоприятные условия (недостаток питательных веществ, воды, большое содержание кислорода, действие высоких и низких температур и т. д.). Спорообразование начинается с появления «спорогенной зоны»: в бактериальной клетке образуется уплотненный участок, где наблюдается обособление ядерного материала и части цитоплазмы с помощью тонкой перегородки. По мере развития и созревания споры закладываются ее стенки, число и толщина которых варьируют у разных видов бактерий (стадия проспоры). Затем проспора уплотняется, уменьшается в объеме, превращается в зрелую спору, которая окружена плотной многослойной оболочкой, состоящей в основном из белков, липидов и гликопептидов. Весь процесс спорообразования длится 18 — 24 ч. По химическому составу споры отличаются высоким содержанием липидов, солей кальция; вода в споре находится в связанном с другими соединениями состоянии. Эти особенности спор и обусловливают их высокую устойчивость к различным факторам: кипячению, действию высоких и низких температур, высушиванию, ультрафиолетовому облучению и т. д. При попадании в благоприятные условия существования (наличие питательных веществ, достаточной влажности и оптимальной температуры) спора прорастает в вегетативную форму: она набухает, в оболочке появляется отверстие, через которое вытягивается росток, превращающийся затем в палочку. Весь процесс длится 4 — 5 ч.
Одной клетке соответствует только одна спора, поэтому спорообразование у бактерий не связано с процессом размножения, как у грибов, а является лишь способом переживания в неблагоприятных условиях внешней среды.
Спорообразующие микробы широко распространены в почве, воздухе, сохраняясь там десятки лет. Среди них встречаются патогенные виды — бациллы сибирской язвы, возбудители газовой гангрены, столбняка и ботулизма.
Сферопласты и протопласты. Бактериальная клетка в определенных условиях может быть лишена клеточной стенки. Эту стенку можно разрушить действием лизоцима или пенициллина, который нарушает синтез гликопептидов. Бактерии, целиком лишенные клеточной стенки, называются протопластами, а при сохранении небольших участков ее — сферопластами. Эти образования покрыты тонкой и нежной цитоплазматической мембраной и имеют сферическую форму. Цитоплазматическая мембрана неспособна сдержать высокое осмотическое давление цитоплазмы, поэтому для сохранения жизнеспособности сферопласты и протопласты помещают в специально осмотически уравновешенные среды, содержащие 5 — 20% сахарозы и сыворотку лошади. В этих средах они сохраняют округлую форму, а некоторые — даже жгутики. Однако такие протопласты неподвижны вследствие нарушения у них механизмов, управляющих движением жгутиков. Спустя некоторое время после хранения сферопластов и протопластов в растворах сахарозы они начинают разрушаться (лизируются) и в среде появляются мелкие зерна и пустые пузырьки — «тени» протопластов. При определенных условиях сферопласты, частично сохраняющие клеточную стенку, могут размножаться на плотных питательных средах и реверсировать (возвращаться) в исходные формы, что сближает их с нестабильными L-формами бактерий типа В.
L-формы бактерий. При частичном или полном разрушении клеточных стенок многие виды бактерий могут образовывать L-формы. Впервые они были обнаружены Клинебергер-Нобель в 1935 г. Название их происходит от первой буквы института Листера (L), в котором они были открыты.
Характерным для L-форм бактерий является их сходство с микроорганизмами группы плевропневмонии крупного рогатого скота (PPLO), которые отнесены в настоящее время к микоплазмам. Однако L-формы отличает от микоплазм то, что им несвойственна потребность в питательных веществах, в которых нуждаются микоплазмы. Генетически L-формы идентичны исходным формам, из которых они получены. У некоторых из них частично сохранена клеточная стенка (L-формы типа В), поэтому они могут превращаться в исходные формы бактерий. Образование L-форм происходит под «действием пенициллина, который нарушает синтез мукопептидов клеточной стенки. Иногда эти формы возникают спонтанно.
По морфологии L-формы разных видов бактерий и других микроорганизмов (трепонемы, дрожжи) сходны между собой. Они представляют шаровидные, вакуолизированные образования величиной от 1 — 8 мкм до мельчайших — 250 нм, способных, как и вирусы, проходить через поры фарфоровых фильтров. Однако в отличие от вирусов L-формы можно выращивать на искусственных питательных средах, добавляя к ним пенициллин, сахара, лошадиную сыворотку. При удалении из такой среды пенициллина L-формы (тип В) вновь превращаются в. исходные формы бактерий. Этот процесс называется реверсией. Однако существуют стабильные L-формы бактерий (тип А), возвращение которых к исходной форме затруднено или невозможно. В настоящее время получены L-формы протея, кишечной палочки, холерного вибриона, бруцелл, возбудителей газовой гангрены, столбняка и других микроорганизмов.
 Занятие 1-е. Вакцины и анатоксины. Вопросы для обсуждения. 1. Искусственный иммунитет, активный и пассивный. 2. Препараты для создания искусственного активного иммунитета: вакцины и анатоксины. 3. Виды вакцин: живые, убитые и химические. 4. Способы приготовления вакцин. 5. Анатоксины нативные и очищенные, их получение и титрован... Читать далее... |
 Занятие 1-е. Методы вирусологических исследований. Вопросы для обсуждения: 1. Особенности биологии вирусов. 2. Принципы классификации вирусов. 3. Вирион, его строение, размеры и химический состав. 4. Микроскопические методы изучения морфологии вирусов. 5. Методы культивирования вирусов на культурах клеток, куриных эмбрионах, лаб... Читать далее... |
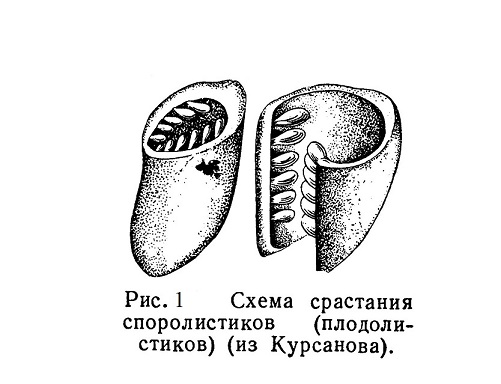 Покрытосеменные, наиболее молодая группа растений, быстро распространились, начиная с середины мезозойской эры, и занимают в настоящее время господствующее положение среди растений земного шара. По сравнению с голосеменными главным новшеством у покрытосеменных является возникновение пестика, образовавшегося из свернувшихся и сросшихся краями плодолистиков.... Читать далее... |
 Чрезвычайно интересную группу представляют открытые в начале текущего столетия так называемые семенные папоротники. Эти растения внешне вполне сходны с папоротниками, но на листьях их найдены настоящие семена с зародышами будущих растений. Дальнейшее изучение показало, что эти растения разноспоровые. Читать далее... |
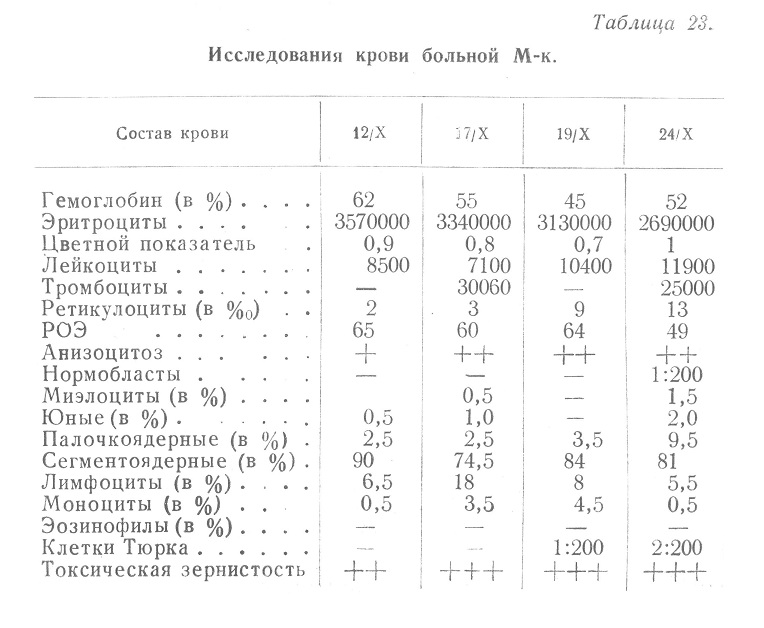 Заслуживают также внимания изменения при пневмонии, осложняющей алиментарную дистрофию. Обращает на себя внимание, прежде всего, противоречивость данных, представленных различными авторами, полученных при очень близких условиях. Так, С. Л. Глухман отмечает, что среди наблюдавшихся им больных лейкопения определялась в 26% случаев, нормальное число лейкоцитов — в 21%, умеренное повышение (8000—12000) в 31% и значительное (12... Читать далее... |
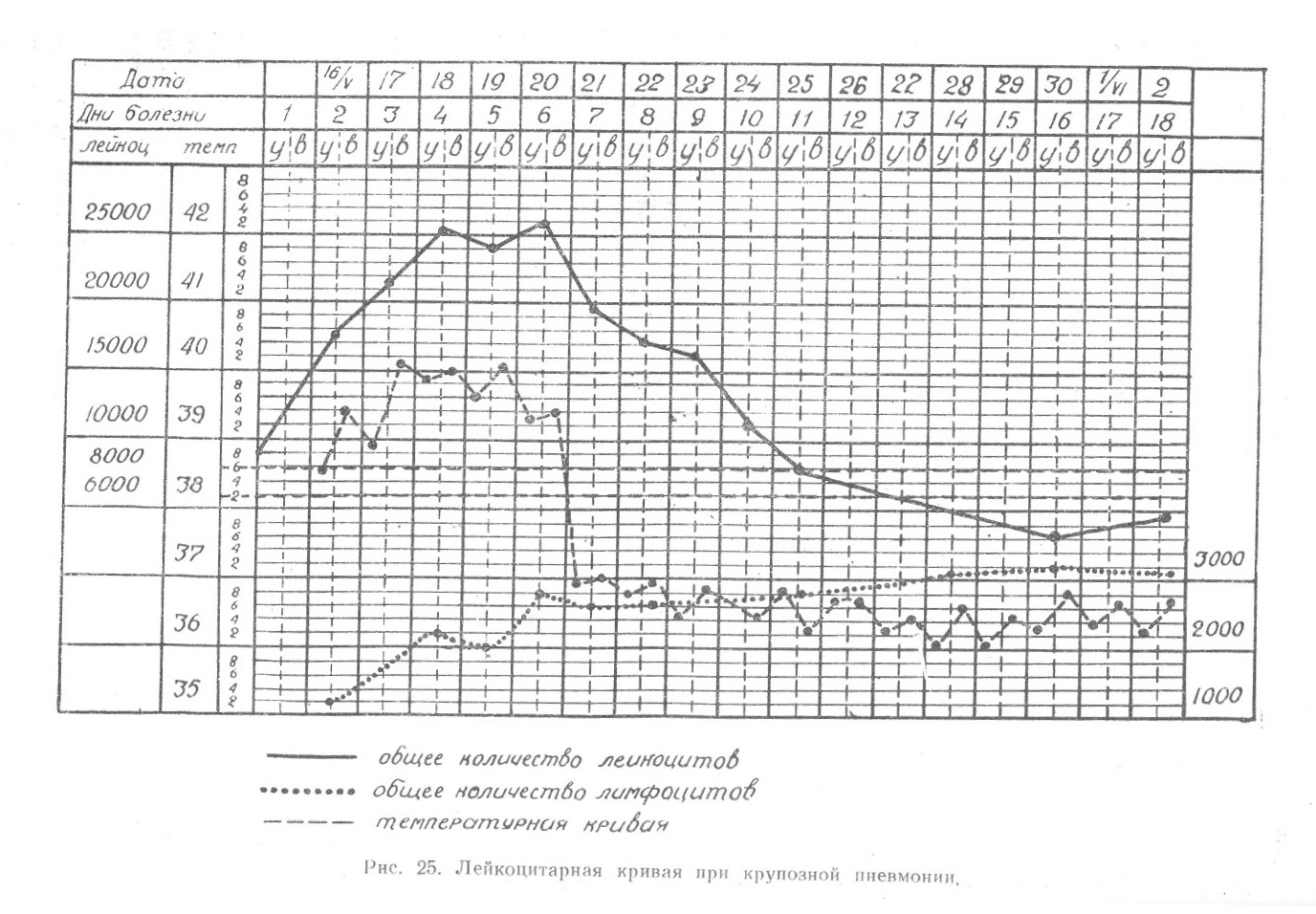 При изложении морфологии мы уже подчеркивали, что ретикулоэндотелиальная система — понятие не только или не столько морфологическое, сколько функциональное. Система эта, ограниченная морфологически (анатомически) ретикулярными и эндотелиальными клетками, многообразна по функциальной деятельности потому, что способна не только к восприятию различных веществ, вводимых извне или образующихся в самом организме в виде грубых взвесей ... Читать далее... |
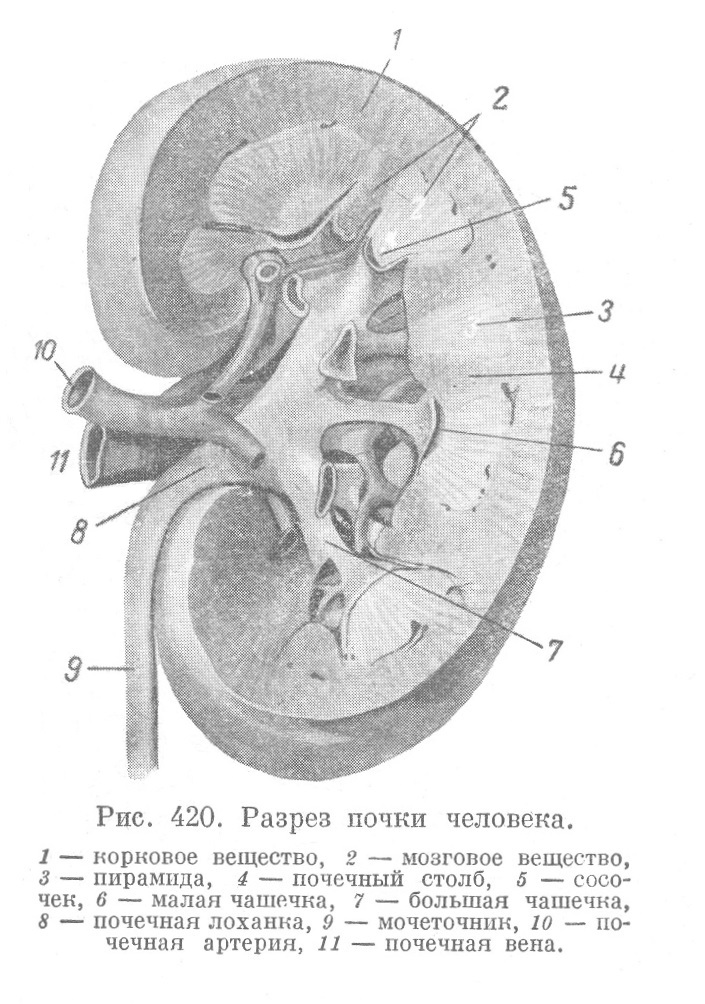 Почки представляют плотный орган бобовидной формы. Средняя часть медиальной (вогнутой) поверхности почки носит название ворот почки (hilus). Отсюда начинается мочеточник и здесь же вступают в почку сосуды (рис. 420). Почка снаружи покрыта легко отделяющейся фиброзной капсулой, состоящей из плотной коллагеновой ткани (с эластическими волокнами), в глубоких частях... Читать далее... |
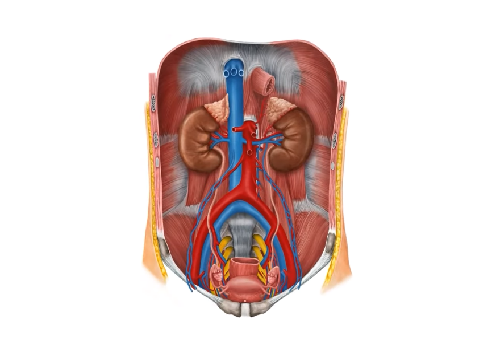 К выделительной системе относятся почки и отходящие от них мочевыводящие пути (малые и большие чашечки, лоханки, мочеточники, мочевой пузырь и мочеиспускательный канал). В почках происходит выделение из кровеносного русла продуктов обмена. Через мочевыводящие пути указанные продукты выводятся из организма. Выделительную функцию в известной степени несут и многие другие органы (кожа, легкие, пищеварительный тракт), однако она не являет... Читать далее... |